filmov
tv
06. Fotorrespiración: Plantas C3, C4 y CAM

Показать описание
La fotorrespiración es un proceso que tiene lugar cuando el ambiente es cálido y seco. En estas condiciones, los estomas de las hojas se cierran para evitar la pérdida de agua y el oxígeno producido durante la fase luminosa de la fotosíntesis alcanza grandes concentraciones. Esto provoca que la Rubisco, la enzima del Ciclo de Calvin que se encarga de unir CO2 a la ribulosa 1,5-bifosfato, se decante por unirse a la molécula 02 y que consecuentemente se produzca la destrucción ribulosa- 1,5- difosfato (5c). Cuando sucede esto hablamos de fotorrespiración y este proceso resulta muy perjudicial porque reduce la capacidad fotosintética de la planta.
Así pues, en el proceso de fotorrespiración, la Rubisco fija O2 en la molécula de 5C, la ribulosa 1,6-bifosfato, produciéndose un compuesto de 3 carbonos, GAP y un compuesto de 2 carbonos, fosfoglicolato. El fosfoglicolato no puede continuar en el ciclo de Calvin y es expulsado.
Ahora bien, para recuperar parte de los carbonos perdidos, las plantas someten 2 moléculas fosfoglicerato a un proceso en el que se acaba produciendo una molécula de PGA que podrá ser reincorporada al ciclo de Calvin, pero se pierde un átomo de carbono en forma de CO2.
Aun así, este proceso no permite la generación de moléculas de glucosa ni mucho menos la regeneración completa de las moléculas de ribulosa 1,5-bifosfato, por este motivo decimos que este proceso disminuye considerablemente el rendimiento fotosintético de las plantas.
Por lo tanto, la fotorrespiración es una vía metabólica derrochadora que sucede cuando la enzima Rubisco del ciclo de Calvin se une a un oxígeno en vez del dióxido de carbono.
La mayoría de las plantas, denominadas C3, no pueden hacer nada para combatir la fotorrespiración. Ahora bien, las plantas C4 y CAM sí que disponen de estrategias para reducir al mínimo la fotorrespiración. Las plantas C4 lo hacen separando la fijación inicial de CO2 y el ciclo de Calvin en espacios separados dado que lo llevan a cabo células de diferentes tipos. Mientras que las plantas CAM separan en el tiempo estos dos procesos.
Empezaremos viendo el funcionamiento de las plantas C4:
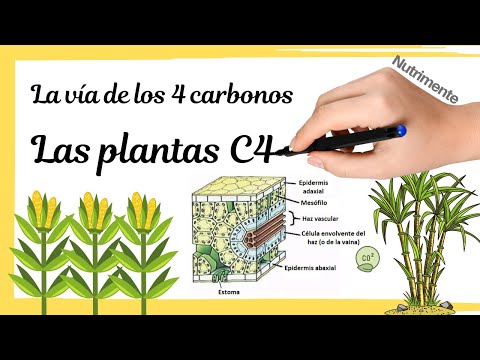
En las células del mesófilo, la enzima PEP carboxilasa, una enzima parecida a la rubisco, pero que no se ve afectada por elevadas concentraciones de oxígeno, capta el CO2 atmosférico y lo une a una molécula de 3C formando una molécula de 4C, el ácido oxalacético.
La molécula en esta forma no puede salir de la célula del mesófilo y por consiguiente es transformada en enfermo, de esta forma será transportada hacia las células internas localizadas en el tejido conductor. Aquí, la molécula de 4, libera una molécula de dióxido de carbono que será captada por la Rubisco e incorporada al ciclo de Calvin permitiendo la generación de materia orgánica, mientras que el piruvato generado deberá devolver a las células del mesófilo consumiéndose 1 molécula de ATP y convirtiéndose en la forma PEP con el fin de que el PEP carboxilasa pueda continuar trabajando ya que necesita este sustrato.
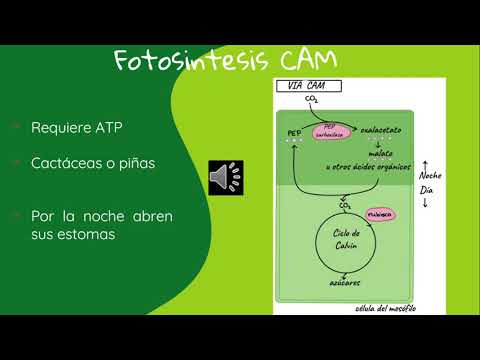
Por lo que respecta a las plantas CAM, estas plantas dominan en zonas muy cálidas y secas como los desiertos. En este caso, estas plantas por la noche, con la oscuridad, abren sus estomas para que el CO2 se difunda por las hojas. Este CO2 se fija al PEP mediante la PEP carboxilasa (lo mismo que sucede con las plantas C4), y el compuesto de 4C se guarda en las vacuolas de las células del mesófilo. Cuando de nuevo la planta dispone de luz, es decir, amanece, el compuesto de 4 carbonos sale de las vacuolas y libera 1 molécula de dióxido de carbono que será utilizada en el ciclo de Calvin para producir materia orgánica.
Recuerdo que la energía y el poder reductor se obtienen de la fase luminosa de la fotosíntesis y que, por lo tanto, sólo se obtienen cuando la planta recibe luz. Por esta razón, el ciclo de Calvin también se produce durante el día, que es cuando se dispone de energía y poder reductor en abundancia.
La vía CAM también consume ATP, sin embargo, las plantas que utilizan esta vía además de evitar la fotorrespiración también utilizan el agua de forma más eficiente, ya que por la noche las temperaturas son más bajas y la humedad es más elevada, y consecuentemente la pérdida de agua por parte de las hojas estas plantas es mucho menor.
Así pues, en el proceso de fotorrespiración, la Rubisco fija O2 en la molécula de 5C, la ribulosa 1,6-bifosfato, produciéndose un compuesto de 3 carbonos, GAP y un compuesto de 2 carbonos, fosfoglicolato. El fosfoglicolato no puede continuar en el ciclo de Calvin y es expulsado.
Ahora bien, para recuperar parte de los carbonos perdidos, las plantas someten 2 moléculas fosfoglicerato a un proceso en el que se acaba produciendo una molécula de PGA que podrá ser reincorporada al ciclo de Calvin, pero se pierde un átomo de carbono en forma de CO2.
Aun así, este proceso no permite la generación de moléculas de glucosa ni mucho menos la regeneración completa de las moléculas de ribulosa 1,5-bifosfato, por este motivo decimos que este proceso disminuye considerablemente el rendimiento fotosintético de las plantas.
Por lo tanto, la fotorrespiración es una vía metabólica derrochadora que sucede cuando la enzima Rubisco del ciclo de Calvin se une a un oxígeno en vez del dióxido de carbono.
La mayoría de las plantas, denominadas C3, no pueden hacer nada para combatir la fotorrespiración. Ahora bien, las plantas C4 y CAM sí que disponen de estrategias para reducir al mínimo la fotorrespiración. Las plantas C4 lo hacen separando la fijación inicial de CO2 y el ciclo de Calvin en espacios separados dado que lo llevan a cabo células de diferentes tipos. Mientras que las plantas CAM separan en el tiempo estos dos procesos.
Empezaremos viendo el funcionamiento de las plantas C4:
En las células del mesófilo, la enzima PEP carboxilasa, una enzima parecida a la rubisco, pero que no se ve afectada por elevadas concentraciones de oxígeno, capta el CO2 atmosférico y lo une a una molécula de 3C formando una molécula de 4C, el ácido oxalacético.
La molécula en esta forma no puede salir de la célula del mesófilo y por consiguiente es transformada en enfermo, de esta forma será transportada hacia las células internas localizadas en el tejido conductor. Aquí, la molécula de 4, libera una molécula de dióxido de carbono que será captada por la Rubisco e incorporada al ciclo de Calvin permitiendo la generación de materia orgánica, mientras que el piruvato generado deberá devolver a las células del mesófilo consumiéndose 1 molécula de ATP y convirtiéndose en la forma PEP con el fin de que el PEP carboxilasa pueda continuar trabajando ya que necesita este sustrato.
Por lo que respecta a las plantas CAM, estas plantas dominan en zonas muy cálidas y secas como los desiertos. En este caso, estas plantas por la noche, con la oscuridad, abren sus estomas para que el CO2 se difunda por las hojas. Este CO2 se fija al PEP mediante la PEP carboxilasa (lo mismo que sucede con las plantas C4), y el compuesto de 4C se guarda en las vacuolas de las células del mesófilo. Cuando de nuevo la planta dispone de luz, es decir, amanece, el compuesto de 4 carbonos sale de las vacuolas y libera 1 molécula de dióxido de carbono que será utilizada en el ciclo de Calvin para producir materia orgánica.
Recuerdo que la energía y el poder reductor se obtienen de la fase luminosa de la fotosíntesis y que, por lo tanto, sólo se obtienen cuando la planta recibe luz. Por esta razón, el ciclo de Calvin también se produce durante el día, que es cuando se dispone de energía y poder reductor en abundancia.
La vía CAM también consume ATP, sin embargo, las plantas que utilizan esta vía además de evitar la fotorrespiración también utilizan el agua de forma más eficiente, ya que por la noche las temperaturas son más bajas y la humedad es más elevada, y consecuentemente la pérdida de agua por parte de las hojas estas plantas es mucho menor.
 0:05:41
0:05:41
06. Fotorrespiración: Plantas C3, C4 y CAM
 0:10:37
0:10:37
Las PLANTAS C4 [La vía de los 4 carbonos] / Comparación con plantas C3
 0:07:32
0:07:32
La Fotorrespiración
 0:06:29
0:06:29
Plantas C3 y C4 - GANADERIA
 0:21:11
0:21:11
Fotosíntesis en plantas C3, C4 y CAM
 0:02:00
0:02:00
¿QUÉ SON LAS PLANTAS C3?
 0:11:30
0:11:30
FOTOSÍNTESIS VÍA C4 - FOTOSÍNTESIS CAM - FOTOSÍNTESIS ANOXIGENICA - FOTORRESPIRACIÓN - BACT. VERDES...
 0:06:14
0:06:14
Las PLANTAS con METABOLISMO CAM
 0:08:00
0:08:00
FOTORRESPIRACIÓN
 0:07:15
0:07:15
Fotosíntesis En Plantas Tipo C3, C4 y CAM
 0:04:18
0:04:18
fotosíntesis plantas C3, C4 y CAM
 0:04:54
0:04:54
Fotosíntesis en plantas C3, C4 y CAM.
 0:05:55
0:05:55
diferencias de las plantas C3, C4 y CAM
 0:17:03
0:17:03
Fotorrespiración
 0:12:44
0:12:44
Fotorrespiración | Fotosíntesis | Biología | Khan Academy en Español
 0:03:58
0:03:58
FOTOSÍNTESIS DE PLANTAS C3,C4 Y CAM 🌄
 0:02:48
0:02:48
🌿Fotosíntesis | Fase Luminosa y Oscura | Ciclo de Calvin🌞 [Fácil y Rápido] | BIOLOGÍA |
 0:03:39
0:03:39
Fotosintesis en Plantas C3, C4 y CAM
 1:12:38
1:12:38
Fotosíntesis: Plantas C3, C4 y CAM
 0:00:58
0:00:58
PLANTAS C3, C4 y CAM 🍍🌵🌴| ¿POR QUE las C4 y CAM están MEJOR ADAPTADAS a Climas Secos☀️?...
 0:02:31
0:02:31
Plantas C3 y C4
 0:02:46
0:02:46
Evaluacion Oral: Diferencia entre fotosíntesis y respiración Planta C3 C4 y CAM
 0:00:36
0:00:36
fotorespiración RUTA DE HATCH SLACK2 * Plantas C3-C4
 0:03:24
0:03:24
Proceso de Fotorrespiración.
Комментарии